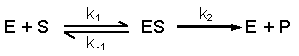Biochemie
Inleiding
In deze module vind je de basisgegevens over de scheikundige processen in levende wezens.
Over de (bio)chemische structuren kun je veel vinden in module 4, maar ook in deze module wordt - waar nodig - een en ander over de structuren van biochemische stoffen (nog eens) besproken.
Het geheel en de samenhang van al die reacties in een levend wezen noemen we ook wel: het metabolisme.
Het gaat vooral over het maken én het afbreken van al die stoffen die een mens, een dier of een plant nodig heeft of juist kwijt wil.
In de biochemie treden heel veel "organische" verbindingen op - het woord zegt het al - van eenvoudig tot buitengewoon ingewikkeld.
Opvallend is dat zo'n 95% van alle stoffen in het menselijk lichaam opgebouwd zijn uit de elementen waterstof, koolstof, zuurstof en stikstof.
Verder is net zo goed opvallend dat datzelfde lichaam voor meer dan de helft eenvoudigweg water is.
De meest specifieke onderwerpen zijn: De sacchariden, de proteďnen, de vetten en de nucleínezuren; ook wel genoemd: koolhydraten, eiwitten, vetten en DNA/RNA.
Een interessante website over inleiding in enkele biochemische onderwerpen vind je
hier
In principe komen we in de biochemie net zulke reacties tegen als in de algemene scheikunde en zeker als in de koolstofchemie. Denk daarbij met name aan zuur-base reacties, redoxreacties, hydrolyse, condensatie en polymerisatie.
Ga nu eerst een paar opdrachten maken die je helpen om in het onderwerp te komen.
Opdracht 1
Salicylzuur reageert met een oplossing van ammoniak. Geef daarvoor de reactievergelijking.
Opdracht 2
Neem onderstaande tekst door en kijk of je die begrijpt. Vervolgens kun je enkele berekeningen maken om de raadgevingen van de dokter te controleren:
Er komt een dame bij de dokter.
"Ik voel me niet goed, dokter. Al dagenlang doodmoe."
De dokter stuurt haar naar het lab voor bloedanalyse. Ze mag over tien dagen terugkomen.
"Ach doktertje, heb je die resultaten al van het lab? Ik blijf maar zo moe."
"Zeker dame, zegt de dokter, ik heb een lijst met gegevens die aantonen dat de pH van je bloed 7,1 is (dat is laag). En je CO2-concentratie is 1,12 mmol/l wat ook al niet normaal is. Nader onderzoek in het academisch ziekenhuis is noodzakelijk. Intussen raad ik u aan om dagelijks bicarbonaat-tabletten te slikken. Waarschijnlijk heeft uw bloed gebrek aan HCO3--ionen.
Kom over tien dagen nog eens terug.
De patient gaat naar huis, begreep er helemaal niets van, maar gaat wel die pillen slikken.
Opdracht 3
Leg de begrippen uit in de volgende tekst:
IJzer komt het menselijk lichaam binnen in de vorm van ionen, via voeding. Het Fe3+ kan zich omzetten in Fe2+ in zuur milieu en met een reductor, bijvoorbeeld ascorbaat. Het Fe2+ komt de pancreas binnen bij een pH van 7 en waar Fe2+ weer Fe3+ wordt. Transferrine transporteert Fe3+; Ferritine slaat de IJzerionen op. Heem bevat Fe2+.
Opdracht 4
Is de volgende bewering waar of onwaar?
"In de spijsvertering ondergaan vele polymeren hydrolyse waarbij ze monomeren vormen."
Inhoud van de module
1. Proteďnen
1.1 Aminozuren; hoe ze reageren
1.2 Productie van proteďnes
1.3 De Ureumcyclus
1.4 Structuren van proteďnes
2. Koolhydraten / Sacchariden / Suikers
2.1 structuren van koolhydraten
2.2 Energie in de Biochemie
2.3 Citroenzuurcyclus (cyclus van Krebs)
3. Vetten / Lipiden
3.1 Vetzuren
3.2 (on)Verzadigd
4. Nucleďnezuren
4.1 De basen van de nucleďnezuren / bouwstenen
4.2 Doorgeven van informatie; genetica
5. Enzymen
5.1 Specificiteit van enzymen
5.2 Structuur van enzymen
5.3 Optimale werking en denaturatie van enzymen
5.4 Regulatie van de enzymactiviteit
5.5 Michaelis Menten
5.6 Bijlage in het Engels over Michaelis Menten
De oude Nederlandse naam voor proteďnen is eiwitten. Je vindt ze in allelevende wezens, dus ook in onszelf.
Ze zijn het belangrijkste bestanddeel van de weefsels van ons lichaam (huid, vlees, nagels, haar, en meer) en worden door het lichaam zelf geproduceerd.
Maar daarvoor zijn wel de bouwstenen nodig: aminozuren. Die moeten van buiten komen via het voedsel, tenminste de meeste daarvan. Sommige aminozuren kan het lichaam zelf bouwen.
Om genoeg aminozuren binnen te krijgen heeft de mens als voedsel vlees, bonen, vis, noten, gelatine en andere producten nodig.
Let op: behalve voor weefsels zijn de proteďnen ook absoluut noodzakelijk in de vorm van enzymen.
Opdracht 5
Is de volgende bewering over aminozuren waar of onwaar?
- aminozuren dienen vooral om vetten te produceren
- Er zijn tien voor de mens essentiële aminozuren
- Alle aminozuren hebben een amfoteer karakter
zuur of base?
Aminozuren bevatten een aminogroep, meestal NH2, en een carboxylgroep, -COOH.
Daarom kan een aminozuur met basen reageren én met zuren. Het aminozuur is een amfolyt.
Als het aminozuur per molecuul 1 aminogroep en 1 carboxylgroep bevat zal de pH van de oplossing van dit aminozuur ongeveer neutraal zijn (6 - 8) Een aminozuur met extra carboxylgroepen zal een lagere pH veroorzaken en een aminozuur met extra aminogroepen zal een hogere pH veroorzaken.
Opdracht 6
Zoek de structuur van Lysine op (zie tabel) en leg uit welke pH een oplossing van Lysine ongeveer zal hebben.
Antwoord 12-06
Het kan heel goed gebeuren dat een aminozuur zich in een omgeving bevindt waar de carboxylgroep zijn H+ afstaat aan een aminogroep.
Dat kan een aminogroep zijn van een ander aminozuurmolecuul, maar soms ook de aminogroep van hetzelfde aminozuurmolecuul. In dat laatste geval is er dus sprake van interne proton-overdracht.
Aan één kant is dat molecuul dan --COO- geworden en aan de andere kant ---NH3+. Eenzelfde molecuul heeft dan dus zowel een positieve alsook een negatieve lading gekregen en we noemen zo'n deeltje ook wel een "dubbel-ion".
Kortom: een aminozuur kan - afhankelijk van het milieu waarin het zich bevindt - op twee manieren neutraal zijn, of positief of negatief zijn.
Opdracht 7
Hoe ziet een eenvoudig aminozuurmolecuul er uit in een omgeving met pH = 7?
en in een omgeving met pH = 2?
en met pH = 10?
Leg je antwoord uit.
Condensatie en Hydrolyse
Behalve in zuur-base reacties kunnen de aminozuren ook prima meedoen aan een ander reactietype: Condensatie. Daarbij wordt de aminogroep van het ene molecuul gekoppeld aan de OH-groep van een ander molecuul, onder afsplitsing van een watermolecuul. Op deze manier worden twee aminozuren aan elkaar gekoppeld d.m.v. een "peptidebinding".
Opdracht 8
Zoek de structuren op van Valine en Alanine (zie tabel) en laat zien hoe deze op twee verschillende manieren in een condensatiereactie aan elkaar kunnen koppelen, waarbij dipeptiden ontstaan.
In scheikundige termen zeggen we: de proteďnen worden gevormd in een "polycondensatieproces", waarbij de aminozuren de "monomeren" zijn.
Je mag proteďnen (eiwitten) beschouwen als een co-polymerisatieproduct omdat er vrijwel altijd verschillende aminozuren meedoen (er zijn zo'n 20 verschillende aminozuren) aan het proces.
Je kunt die hele synthese van eiwitten ook nog op een andere manier bekijken, namelijk als een proces dat gecontroleerd wordt door DNA/RNA. Dat onderwerp behandelen we verderop.
In het volgende schema zie je de deelnemende aminozuren als dubbel-ionen, wat het condensatieproces in het geheel niet beďnvloedt.
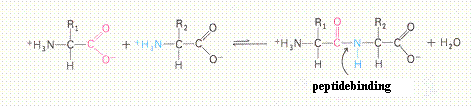

Aan de twee uiteinden van zo'n dipeptidebinding bevinden zich nog steeds: een amino groep en een carboxylgroep. Oftewel, de mogelijkheid om door te gaan blijft aanwezig.
Opnieuw kunnen die groepen 'condenseren' met andere aminozuren en tri-, tetra-, oligo- of polypeptiden vormen.
Een eiwit (proteďne) kan bestaan uit 50 tot 2000 aan elkaar gekoppelde aminozuren en de molecuulmassa varieert van 5.000 tot 20.000
Een paar oefeningen ter herinnering:
Opdracht 9
- Welke zijn de essentiële aminozuren?
- Waarom worden ze zo genoemd?
Optische isomerie
In de natuur heb je maar één soort aminozuren, dat zijn de α-aminozuren. Dit heeft alles te maken met het onderwerp "optische isomerie".
Alle aminozuren die in de natuur voorkomen hebben een aminogroep in de α-positie t.o.v. de carboxylgroep.
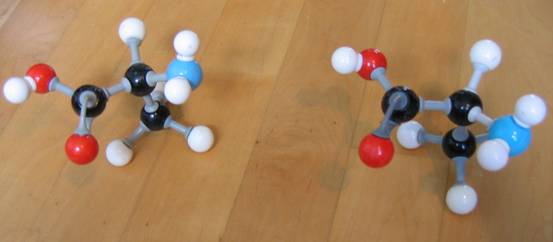

In de afbeeldingen zien we Alanine. Boven α-alanine (twee keer; de twee structuren zijn gelijk en ze zijn niet elkaars spiegelbeeld).
Onder zie je α-alanine en β-alanina, en deze twee zijn optische isomeren (ze zijn niet idenktiek in driedimensionale zin, maar de twee zijn wel elkaars spiegelbeeld).
Opdracht 10
Geef van Alanine de vier verschillende structuren (met en zonder ladingen)
Antwoord 12-10
Hieronder zie je de structuur van penicilline (twee keer).
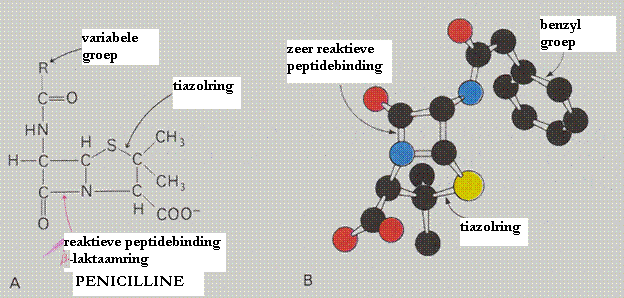
Let op de belangrijke functionele groepen: peptide en carboxyl en het thiazol (waar S een O vervangt).
Opdracht 11
Waar of onwaar?
- De vorming van eiwitten onder controle van DNA is een polymerisatie.
- De vorming van eiwitten onder controle van DNA is een polyhydrolyse.
Er bestaat een aminozuur (Cysteďne) dat een ander soort binding kan maken, namelijk een zwavelbrug (S-S).
Als een heleboel aminozuren een polypeptide maken, dan zal de lange keten zich gaan opvouwen / oprollen tot bepaalde structuren.
In de eerste plaats vormt zich een zgn. Helix, een schroefstructuur, die we de secundaire structuur van eiwitten noemen.
De primaire structuur is eenvoudigweg de volgorde van de aminozuren.
De Helix kan - op zijn beurt - zich weer opvouwen tot een driedimensionale structuur, die we dan de tertiaire eiwitstructuur noemen. (zie figuur)
Hoofddoel van deze cyclus is de vorming van ureum, een belangrijk eindproduct dat als afvalstof wordt uitgescheiden uit het lichaam. Via deze cyclus raakt het lichaam - beetje bij beetje - overtollig stikstof kwijt.
Normaal gesproken komt stikstof het lichaam in via aminozuren.
Deze cyclus is verbonden met de Krebs-cyclus (verbranding van suiker in vele stappen) en deze samenwerking levert een metabolisme op (een sterk katabolisme, een stivige afbraak van grote moleculen in het lichaam).
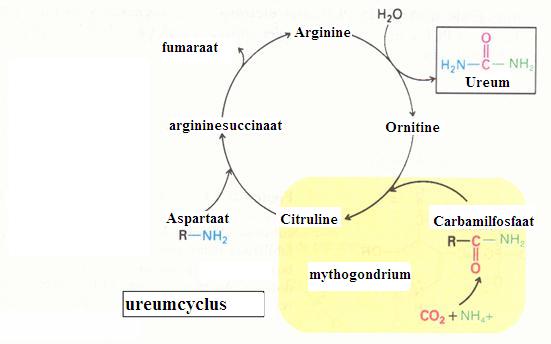
De ureumcyclus dient tegelijk ook voor de afbraak van afgewerkt aminozuur.
Het afvalproduct ureum verlaat het lichaam via de urine.
Opdracht 12
Kijk goed naar de afbeelding en probeer de secundaire en de tertiaire structuur te zien / aan te wijzen én de actieve plek (het co-enzym) van het enzym.
De L-aminozuren zijn de bouwstenen van de eiwitten; zij vormen de peptidebindingen en zijn verantwoordelijk voor de Helixen.
De primaire structuur van de eiwitten
De volgorde van aminozuren in de polypeptiden, in de proteďnen, noemen we dus de primaire structuur. Het is de lineaire volgorde van de aminozuren.
Deze volgorde is buitengewoon belangrijk voor het functioneren van dat eiwit. Vaak zal het vervangen van één aminozuur genoeg zijn om de activiteit van het eiwit in het organisme flink te veranderen, misschien wel met zeer negatieve gevolgen.
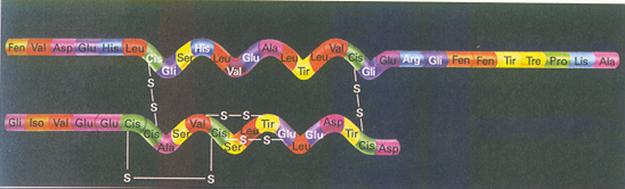
figuur: albumine, De volgorde van de aminozuren en de plaatsen van de zwavelbruggen.
Een eiwitmolecuul kan samengesteld worden uit één of meerdere polypeptiden.
Voorbeeld:
Het albumine (hoofdbestanddeel van eiwit van een ei) heeft één enkele opgerolde polypeptidestructuur,.
Insuline (zie wikipedia) echter, het hormoon dat in de pancreas wordt gemaakt en het gehalte aan glucose in het bloed controleert, wordt samengesteld uit twee polypeptidestructuren.
Opdracht 13
Stel je een eiwitstructuur voor met 1000 monomeren van het aminozuur Valine (zie tabel).
Bereken de molecuulmassa.
Antwoord 12-13
Het aminozuur Proline dat ergens opduikt in een polypeptideketen verbiedt of verstoort de vorming van de α-helixen. Gevolg: er kunnen geen waterstofbruggen worden gevormd (wegens gebrek aan de juiste polariteit).
Voorbeeld:
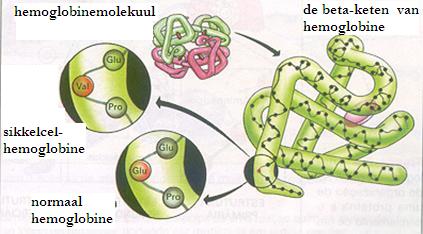
Sikkelcelanemie is een erfelijke anemie (iets wat ontbreekt) en wordt veroorzaakt doordat één bepaald aminozuur ergens in de polypeptideketen van hemoglobine vervangen is door iets anders. (zie ook figuur) Het juiste aminozuur ontbreekt.
De primaire structuur van de proteďnen
is eenvoudigweg de volgorde van de aminozuren. (die is bij cikkelcelanemie dus niet goed).
De secundaire structuur van de proteďnen
De polypeptideketens krullen zich op volgens een soort schroefdraad; we spreken ook wel van een "helix". Dat noemen we de secundaire structuur van het eiwit.
Tertiaire structuur van proteďnen
Een lange polypeptideketen die al een helix gevormd heeft, dan nog steeds zo lang zijn dat die helix zich gaat opvouwen op een driedimensionale manier: de tertiaire structuur.
Quaternaire structuur van proteďnen
De zeer grote eiwitmoleculen zijn samengesteld uit meerdere tertiaire bollen: quaternaire structuur.
Voorbeeld:
In het geval van hemoglobine van het bloed worden vier polypeptideketens, die elk al een tertiaire structuur hadden gevormd, samengevoegd tot één groot geheel: de quaternaire structuur.
De krachten die de vorming van secundaire en tertiaire structuren mogelijk maken zijn:
- Waterstofbruggen
- vanderWaals krachten
- dipolaire krachten
- Zwavelbruggen (Cisteíne)
Opdracht 14
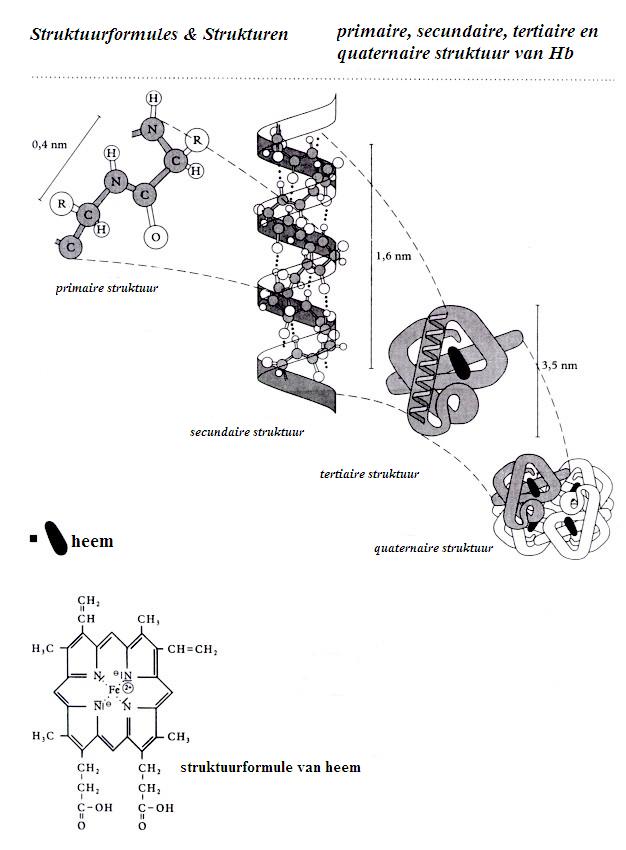
Bestudeer goed bovenstaand diagram van hemoglobine en beantwoord de volgende vragen:
- wat betekent 'nm'?
- Waar in het enzym bevindt zich de helix?
- Wat voor krachten stabilizeren de helix (aangegeven met puntjes . . . . . . . )?
- Wijs het co-enzym aan. waar zit dat?
- Heeft het Hb ergens een anorganisch gedeeltje?
- Wat voor krachten stabilizeren de tertiaire structuur?
Opdracht 15
Waar of onwaar?
- Proteďnen zijn polymeren;
- Proteďnen hebben een primaire structuur, dat is de volgorde van de aminozuren;
- Een secundaire structuur verbindt diverse primaire structuren met elkaar;
- De tertiaire structuur is de helixschroefstructuur;
- De secundaire en tertiaire structuur danken hun stabiliteit aan waterstof- en zwavel-bruggen/bindingen;
- Proteďnen kunnen gedenatureerd worden (reversibel of irreversibel), wat betekent dat ze volledig worden kapotgemaakt met alcohol, met zuur of door verwarming;
- De primaire structuur gaat bij denatureren niet kapot. Dat kan alleen in een chemisch proces (hydrolyse
 aminozuren)
aminozuren)
- Er bestaan "structuurproteďnen" (zoals albumine) en enzymproteďnen (zoals oxidase)
Even ter herinnering: de Indeling van de proteďnen
Je kunt ze indelen naar vorm en dan heb je eiwitten in vezelvorm en in globulaire vorm.
- Vezeleiwitten - toch zeker 10 x zo lang als breed. Zo heb je bijvoorbeeld in de nagels, in huid en huiden: Keratine.
- Globulaire proteďnen - zijn veel ronder van vorm, dus lengte en breedte verschillen niet veel. Voorbeelden: enzymen, antilichamen, membraanproteďnen, hemoglobine, chlorofyl, en meer.
Deze indeling heeft direct te maken met de twee belangrijkste functies van proteďnen:
- vorming van weefsel (spieren, huid, been, colageen, enz.)
- katalyse (controle van de reacties, transmissie van zenuwimpulsen, immuunbescherming)
De studie over proteďnen vormt de basis voor het begrijpen van de genetica.
In het laboratorium zal t.b.v. zulke studies in elk geval nodig zijn: zuivering en scheiding van proteďnen met technieken zoals chromatografie en elektroforese.
Letten we op de molecuulsamenstelling van de eiwitten, kun je de proteďnen indelen in: eenvoudig en complex.
- Eenvoudige proteďnen of homoproteďnen: - zijn uitsluitend opgebouwd uit aminozuren en niks anders. Voorbeelden: insuline, albumine, keratine, fibrinogeen (dat zich bevindt in het bloedplasma en dat meedoet aan de bloedstolling).
- Complexe eiwitten of heteroproteďnen: - zijn gekoppeld met niet-eiwitstructuren, zoals de prostetische groepen.
Opdracht 16
Hoe klassificeren we de proteďnen van vlees? (tot welk soort proteďnen behoren zij?)
Voorbeelden van groep B:
- Chlorofyl, waarvan de prostetische groep Mg is
- Hemoglobine waarvan Heem met Fe de prostetische groep is
- kaseíne (in melk), waarvan de prostetische groep fosfaat is
- kerneiwitten: - voornamelijk in de ribosomen en waarvan de prostetische groep een ribonucleínezuur is (RNA) of (DNA)
- glycoproteďnen, waarvan de prostetische groep een saccharide is; lipoproteďnen met lipiden als prostetische groep
N.B.
- Complexe proteďnen worden ingedeeld naar hun prostetische groep.
- Als de prostetische groep een bepaalde kleur heeft, wordt ook wel gesproken van chromoproteďnen.
Wat betreft hun fundamentele functie worden de proteďnen ingedeeld in structuureiwitten en enzymen:
- structuureiwitten:
behoren bij de celstructuur en zijn daar "bouwstoffen"
- Enzymeiwitten:
controleren praktisch alle chemische reacties in de cellen.
Opdracht 17
Hemoglobine dankt zijn rode kleur aan de heemgroep.
Probeer nu hemoglobine te klassificeren en wat is het meest karakteristiek voor de heemgroep?
Tenslotte: eiwitten kun je 'denatureren'.
D.w.z.: het molecuul wordt verwarmd tot bij de negentig graden Celcius. De quaternaire, de tertiaire en zelfs de secundaire structuur gaan dan geheel of gedeeltelijk verloren. Je kan zeggen dat het resultaat is: een losse lange draad van aminozuren. De primaire structuur is als enige behouden.
Dikke kans dat bij afkoelen een goed deel van die moleculen weer hun oude structuren terugkrijgen.
(zie ook wikipedia)
Koolhydraten, ook sachariden genoemd, zijn een bepaald type verbindingen van koolstof-, waterstof- en zuurstofatomen waarbij de waterstof- en zuurstofatomen in een verhouding 2:1 voorkomen: algemene formule Cn(H2O)m. Ze kunnen oppervlakkig als een hydraat van koolstof worden beschouwd.
Opdracht 18
Is de volgende bewering waar of onwaar? Leg uit.
"De naam 'Koolhydraten' baseert zich op een oud wetenschappelijk misverstand."
Lang niet alle verbindingen in de levende natuur die als koolhydraten zijn geclassificeerd beantwoorden evenwel aan deze verhoudingsformule. Ook derivaten met andere atoomtypen en andere verhoudingen worden tot de omvangrijke groep van de koolhydraten gerekend zolang ze dezelfde basisstructuur hebben. Zo worden ook desoxyribose (C5H10O4) en glucosamine (C6H13O5N) tot de koolhydraten gerekend, melkzuur (C3H6O3) daarentegen niet.
Koolhydraten zijn leveranciers van de voor het leven benodigde energie en vormen als zodanig het belangrijkste voedingsbestanddeel van vele dieren en de mens. In planten worden zij door fotosynthese gevormd uit kooldioxide en water, in andere organismen ontstaan zij uit wat complexere koolstofverbindingen van ten minste twee of drie koolstofatomen. In de stofwisselingsprocessen van levende organismen nemen de afbraak en synthese van koolhydraten een centrale plaats in.
Suiker is in het dagelijks gebruik de naam van een zoetsmakende chemische verbinding die veel in de keuken wordt gebruikt. Deze verbinding wordt beschreven onder sacharose.
Eenvoudige indeling van de sachariden:
- Monosachariden: glucose, fructose, galactose(C6H12O6)
- Disachariden: saccharose (suiker), lactose, maltose (C12H22O11)
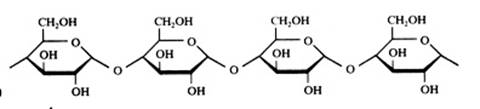
- polysachariden: amide (zetmeel), amilose, cellulose( (C6H10O5)n)
Structuren:
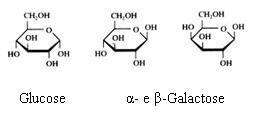
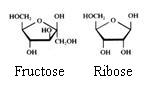
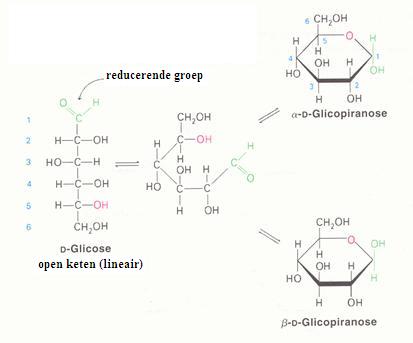
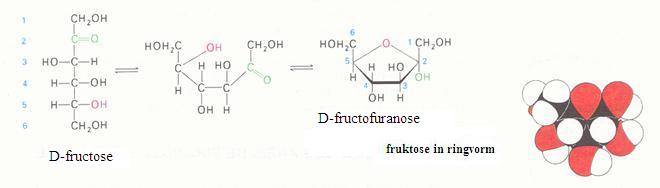
Monosachariden
De monosachariden kunnen zich bevinden in lineaire of in cyclische vorm (hierboven zie je alleen cyclische vormen).
Je kunt in onderstaande schema zien hoe glucose de lineaire en de cyclische vorm kan aannemen:
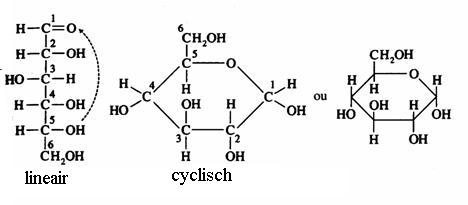
De lineaire structuur kan gemakkelijk geoxideerd worden, dus met een zwakkere oxidator; vaak gebruikt men:
Ag+ ammoniakale zilveroplossing = reagens van Tollens. Ook Fehlings' reagens werkt prima (Cu2+)
De cyclische vorm wordt niet gemakkelijk geoxideerd (er is geen =O binding, geen aldehydegroep, beschikbaar).
Het is in de praktijk nu eenmaal zo dat de twee structuren, de lineaire en de cyclische in oplossing met elkaar in evenwicht zijn, dus dat de lineaire structuur er altijd bij zit en die laat zich goed oxideren.
Omdat de lineaire tijdens dat proces verdwijnt en in evenwicht is met de cyclische, zal de cyclische vorm steeds meer overgaan in lineaire vorm, oftewel, alles wordt uiteindelijk toch geoxideerd.
Opdracht 19
Bestudeer goed de volgende twee afbeeldingen: glucose en fructose komen op twee manieren voor, de lineaire en de cyclische. De twee vormen zijn - in watermilieu, altijd in evenwicht met elkaar.
Toch heeft alleen de lineaire structuur de juiste groep om te oxideren (de reductorgroep).
Leg met eigen woorden uit waarom tijdens het redox-proces toch niet alleen de lineaire vorm, maar ook de cyclische vorm verdwijnt.
Disacchariden
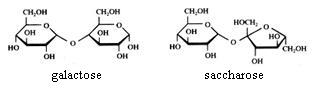
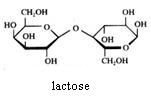
Saccharose kan niet geoxideerd worden met Ag+ of met Cu2+.
Maltose en lactose kunnen wel geoxideerd worden met Ag+ of met Cu2+.
Amide, zetmeel wordt niet geoxideerd met Ag+ of met Cu2+.
Opdracht 20
De oxideerbaarheid van een cyclische structuur hangt af van de plaats van de O (zuurstofatoom) in de ring.
Als zo'n C - O binding in de ring gemakkelijk is te openen (waarbij de lineaire structuur ontstaat) dan is oxidatie mogelijk.
(zie ook module 11)
Analyseer, met behulp van de structuren, waarom sommige ringen wel en ander niet gemakkelijk open gaan.

Polysacchariden
Amide of zetmeel heeft een helix-structuur als het zich in natuurlijke vorm bevindt.
In dat geval maakt het met I3--ionen een donkerblauw gekleurd complex.
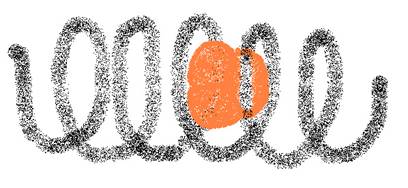
Opdracht 21
Jodium is een tamelijk apolaire stof en lost normaal niet op in water. Maar wel als dat water ook jodiumionen bevat, bijvoorbeeld van KI.
Leg dat uit.
Fehlings' Reagens
CuSO4(aq) + 2NaOH(aq)  Cu(OH)2(s) + Na2SO4(aq)
Cu(OH)2(s) + Na2SO4(aq)
Cu(OH)2(s) Cu2+ + 2OH-( = evenwicht 1)
Cu2+ + e- Cu+
De toevoeging van NatriumKaliumTartraat: de tartraat-ionen grijpen de Cu2+-ionen van evenwicht 1 waardoor het neerslag Cu(OH)2 verdwijnt; tegelijk verschijnt er een transparante oplossing met donkerblauwe kleur = Fehlings' Reagens (FR).
FR bezit dus in basisch milieu opgeloste Cu2+-ionen die een reductor kunnen oxideren (bijvoorbeeld de aldehydegroep), door een elektron te pakken en een ion Cu+ te vormen.
Deze ionen Cu+ zullen in een vervolgreactie met OH- het oranjerode neerslag Cu2O(s) vormen.
Dit wordt wel gebruikt als een bewijsreactie voor de aanwezigheid van monosacchariden.
Opdracht 22
Waar of Onwaar?
Om de aanwezigheid van glucose in urine te onderzoeken is een specifieker methode dan Fehlings' Reagens de enzymatische methode met glucose-oxidase.
Antwoord 12-22
Aspartaam is een stof die wordt gebruikt door mensen die willen afvallen.
Ze willen voorkomen dat ze vet worden van te veel suiker of ze mogen om andere redenen geen suiker hebben.
Het is nu eenmaal zo dat alle suikers nogal veel energie bevatten. In plaats van suiker in de koffie gebruikt men dan vervangers zoals aspartaam, maar er zijn ook andere.
Veel suikerhoudende producten (bijvoorbeeld jam en coca-cola) hebben ook zogenaamde dieetproducten, d.w.z. zonder suiker, maar met vervangende zoetstoffen.
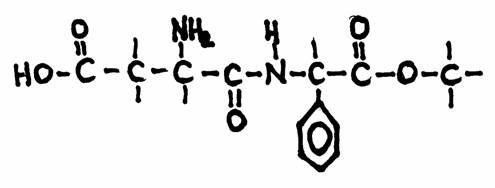
Het geheim is dat aspartaam het lichaam in gaat zonder aan het metabolisme mee te doen, zonder enige energie-waarde, zonder enige vetvorming.
Maar een waarschuwing is wel nodig: sommige suikervervangers blijken inmiddels, als je ze erg veel gebruikt, niet zo gezond omdat ze kanker kunnen veroorzaken.
Opdracht 23
Bestudeer de aspartaamstructuur nog eens goed en wijs daarin aan wat je kent, de functionele groepen dus.
In principe zijn de regels voor exotherme en endotherme reacties in de biochemie gelijk aan de regels voor de scheikunde in het algemeen. Die gelden dus ook voor de chemische reacties in levende wezens. Ook daar vinden chemische reacties plaats die energie opleveren of energie kosten.
Wel is het zo dat de energie in levende wezens vaak gedragen en overgebracht wordt via speciale stoffen, zoals ATP (Adenosine Tri Phosphaat), een molecuul dat vaak om de hoek komt kijken bij reacties die gepaard gaan met energieoverdracht.
De meeste levende wezens moeten ademhalen, waarbij het gaat om het binnenkrijgen van zuurstof (respiratie).
Groene planten hebben nóg een mogelijkheid: de fotosynthese die ongeveer het tegenovergestelde is van ademhaling. Bij de fotosynthese wordt zuurstof gemaakt in plaats van verbruikt.
Respiratie en fotosynthese zijn niet de enige biochemische processen met stevig energie-effect; in feite zijn ALLE biochemische reacties exotherm of endotherm, maar daarover gaan we het hier niet hebben.
Fotosynthese en ademhaling
Dat zijn twee tegengestelde processen in de natuur:
- de respiratie: C6H12O6 + 6O2 6CO2 + 6H2O exotherm(ΔH < 0)
- de fotosinthese: 6 CO2 + 6H2O C6H12O6 + 6O2 endotherm (ΔH > 0)
Niet alleen mensen en dieren, maar ook planten "ademen"!
In tegenstelling tot de fotosynthese, geldt voor de ademhaling dat er chemische energie van buiten (in de vorm van zuurstof en bijvoorbeeld suikers) moet worden toegevoerd.
De fotosynthese daarentegen produceert juist chemische energie.(bijvoorbeeld ATP en de suikers die bij de ademhaling weer worden verbruikt).
Planten hebben dus beide mogelijkheden, overdag domineert de fotosynthese (o.i.v. licht) en 's nachts de ademhaling.
Opdracht 24
Stel je eens voor dat ergens een mens er in geslaagd is (bijvoorbeeld door mutatie van zijn genen) de mogelijkheid te verkrijgen om ergens in zijn lichaam fotosynthese te realiseren.
Wat zouden de consequenties zijn?
Antwoord 12-24
Er is nog iets anders om over te dromen:
Stel dat we - in plaats van de bekende zonnecellen in zonnepanelen - chlorofylmoleculen kunnen maken en in grote hoeveelheden op zo'n zonnepaneel konden plaatsen. Dan zou op de meest effectieve manier zonne-energie omgezet kunnen worden in waterstof of zelfs direct in elektriciteit.
Dat zou nog eens een revolutie zijn op de energiemarkt. Het schijnt dat er hier en daar onderzoek naar gedaan wordt.
Net als de Ureumcyclus, vindt ook de Krebscyclus plaats op de mithogondria (de energiefabriekjes in de cellen).
De oxidator (zuurstof, O2) helpt bij de 'decarboxylering' van het citroenzuur, tijdens het verloop van al die reacties van deze cyclus.
Eén complete omgang door de cyclus houdt een reductie in met twee koolstofatomen die overgaan zijn 2CO2).
De bijbehorende waterstofatomen verbinden zich aan energiedragers: FAD en NAD, die meteen daarna ATP vormen. ATP mogen we beschouwen als een soort brandstof in de levende wezens.
De ATP-moleculen zitten vol chemische energie.
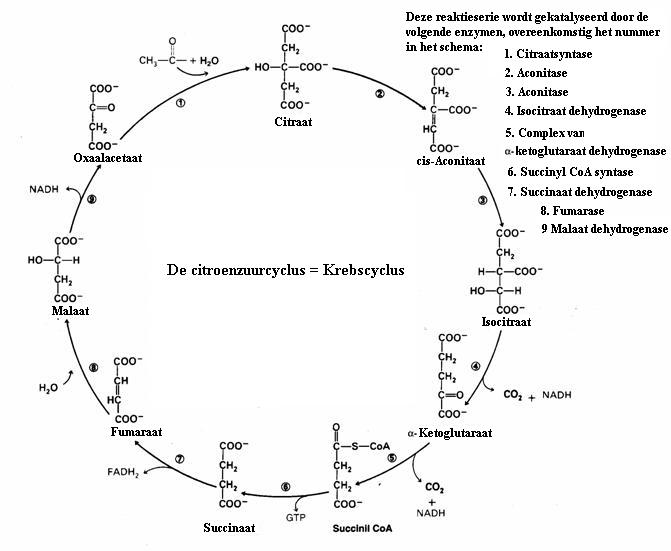
De hele cyclus is opgebouwd uit negen stappen, deelreacties, met als totaalreactie:
| Citraat
|
|
|
|
oxaal acetaat
|
|
| C6H5O72-
|
+
|
H2O
|
|
C4H2O52-
|
+ 2CO2 + 5H·
|
FAD- en NAD-moleculen pakken de H-radikalen op (neutrale eenzame H-atomen) en verkrijgen daarmee flink veel energie.
Daarna reageren deze H-atomen met zuurstof en vormen water H2O (zeer exotherme reactie).
Water en koolzuurgas zijn de eindproducten.
De hoofdfunctie van de cyclus is het verkrijgen en opslaan van chemische energie.
Opdracht 25
Wat kun je zeggen over de elektronenoverdracht in bovengenoemde processen?
In de natuur komen we vetten vooral tegen in dierlijk vet en plantaardige olie. Die twee kunnen heel goed dienen als voedsel voor mens en dier.
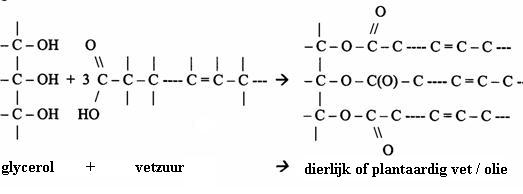
Ze vormen zich in een condensatieproces van gycerol met vetzuren. Glycerol heeft drie OH-groepen en als die alledrie meedoen aan het condensatieproces heten de producten: "triglyceriden".
Vetzuren hebben een chemische structuur met uiteraard een carboxylgroep (het zijn zuren immers) + een koolstofketen van zo'n 15 tot 20 C's.
Meestal komen de vetzuren voor in verbinding met glycerine, maar hier en daar komen ze ook vrij voor (bijvoorbeeld, stearinezuur = kaarsvet = , C17H35COOH).
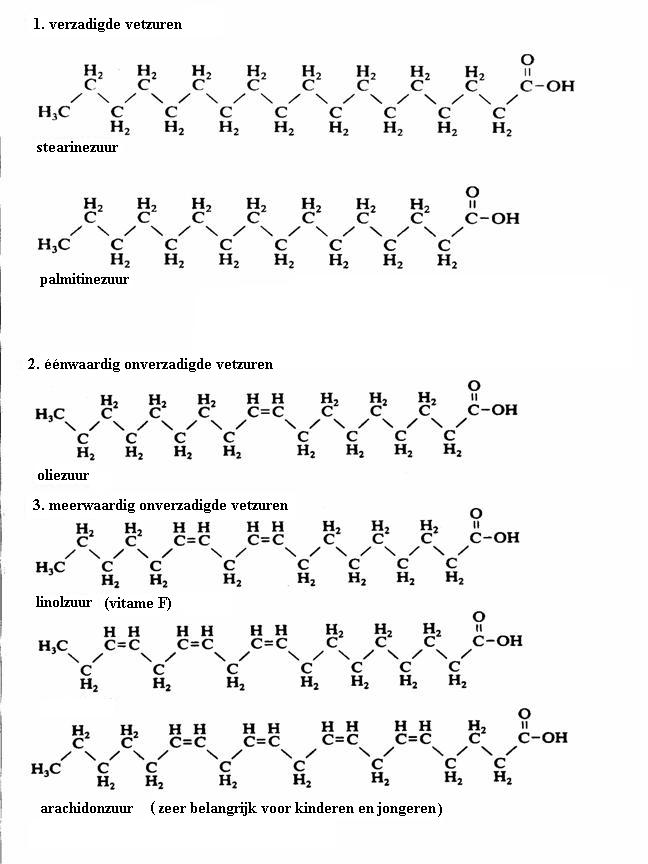
In de figuur zie je de structuren van drie vetzuren.
Eén daarvan in stearinezuur (kaarsvet); de ander heeft een dubbele binding, dus is onverzadigd; de derde heeft meerdere dubbele bindingen, dus is meervoudig onverzadigd.
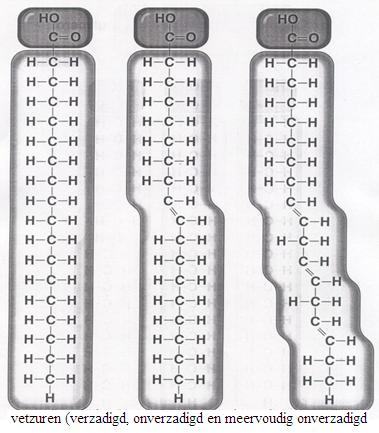
De volgende afbeelding toont vier modellen: drie modellen van onverzadigd vetzuur + één model van glycerine.
De drie vetzuren gaan reageren, elk met één OH-groep van het glycerine, onder gelijktijdige vorming van drie moleculen water.

En in de volgende afbeelding kun je de producten zien: een vetmolecuul + de drie gevormde watermoleculen:

In de natuur zijn de lipiden (vetten) de mono-, di- en tri-glyceriden van vetzuren, of ook: esters van glycerol en vetzuren.
"Vet" is verzadigd.
Gebruiken we onverzadigde vetzuren: dan wordt het vette karakter minder en het olie-karakter meer.
Olie-achtige stoffen zullen over het algemeen onverzadigd zijn en vette stoffen verzadigd.
Onverzadigde stoffen hebben één of meer dubbele bindingen in de keten die verzadigd kunnen worden, bijvoorbeeld met Broom (Br2)
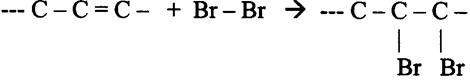
Een laag met onverzadigde olie (gele kleur) laag met vet (troebel)
Een laag met Br2(aq) (gele kleur) laag van water zonder broom
(kleurloos)
Opdracht 26
Waar of Onwaar?:
De functie van gal kun je vergelijking met de functie van zeep.
Opdracht 27
Leg alle begrippen uit van de volgende tekst:
"Normaal gesproken wordt margarine gemaakt uit de grondstof 'plantaardige olie'. Die olie is vooral opgebouwd uit onverzadigde moleculen.
Het probleem zal zijn dat olie een vloeistof is en hoe moet je dat als boter op je brood smeren?
Je kunt dat oplossen (wat men vroeger deed) door de dubbele bindingen te verzadigen met waterstof (katalysator:Zink). Meer verzadigd betekent: harder, dus meer een vetkarakter (smeerbaarder).
Maar tegelijk verliest de stof zijn gezonde karakter.
De nieuwe manier om van die vloeistof een vaste (en smeerbare) stof te maken is de toepassing van emulgators.
Onderstaande inleiding komt grotendeels uit wikipedia.
Hieronder een sterk vereenvoudigd model van een dubbelstrengig nucleďnezuur.
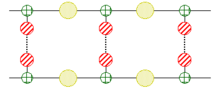
Geel = fosfaat; groen = pentose; rood = stikstofhoudende base.
De doorgaande lijnen vertegenwoordigen de covalente bindingen en de onderbroken lijnen de waterstofbruggen.
Een nucleďnezuur of kernzuur is een complex biochemisch macromolecuul waarin een groot aantal bouwstenen, de nucleotiden, aan elkaar geschakeld zijn. Nucleďnezuren hebben een hoog molecuulgewicht. De volgorde van de nucleotiden bepaalt de genetische informatie.
De meest voorkomende natuurlijke nucleďnezuren zijn desoxyribonucleďnezuur (DNA) en ribonucleďnezuur (RNA).
De naam 'nucleďnezuur' werd gegeven omdat deze moleculen het eerst ontdekt werden in de celkern (nucleus).
Ribonucleďnezuur
Ribonucleďnezuur of RNA is een nucleďnezuur waarin de suiker ribose voorkomt. Het speelt een grote rol bij het overbrengen van genetische informatie van het desoxyribonucleďnezuur (DNA) naar de ribosomen. Op basis van de genetische informatie kunnen proteďnen gemaakt worden.
Er zijn drie types van RNA afhankelijk van hun functie: tRNA (transfer), mRNA (messenger) en rRNA (ribosomaal).
Desoxyribonucleďnezuur
In desoxyribonucleďnezuur vervult de suiker desoxyribose de functie van ribose in RNA. DNA komt voor in alle levende organismen en bevat de erfelijke informatie van het organisme. De DNA-segmenten die genetische informatie bevatten worden genen genoemd. Andere delen hebben een structureel doel of spelen een rol bij de transcriptie van de genetische informatie.
Er zijn 4 + 1 basen, basisstoffen, bouwstenen voor de nucleďnezuren, die het Desoxi ribo nucleďnezuur (DNA) en het Oxiribonucleďnezuur (RNA) construeren, samenstellen.
- DNA wordt opgebouwd door en is dus samengesteld uit Adenine, Guanine, Thymine en Cytosine (A,G,T,C)
- RNA: Adenine, Guanine, Uracil en Cytosine (A,G,U,C)
- Het verschil tussen Thymine en Uracil is slechts één methylgroep.
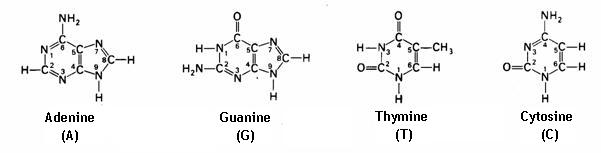
Nucleďnebasen zijn complementair en vormen basenparen.
Volgende paren zijn mogelijk:
- cytosine-guanine (C-G) (bij DNA en RNA)
- adenine-thymine (A-T) (bij DNA)
- adenine-uracil (A-U) (bij RNA)
De binding tussen cytosine en guanine heeft drie waterstofbruggen, terwijl het A-T-paar twee waterstofbruggen vormt.
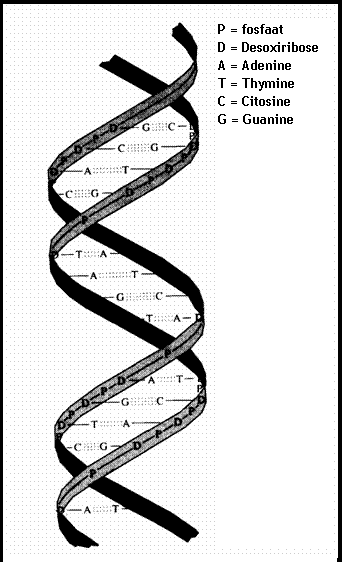
De dubbele helix van DNA, schematisch en in model:
In zo'n (super)macromolecuul van DNA gaan twee zeer lange helixen samen (zie figuur). Altijd is er die aantrekking tussen G en C of A en T.
"DNA"

Nucleosiden
Als een van de vijf basen zich koppelt aan een suiker(ring), heet dat een nucleoside.
Nucleotiden
Wanneer een fosfaatgroep aan een nucleoside wordt toegevoegd, krijg je een nucleotide.
Nucleotiden zijn de monomeren van DNA en RNA.
Opdracht 28
Binnen de dubbele helix nemen de moleculen A, G, T, C e U deel aan de bindingen tussen de twee strengen van die dubbele helix. (zie figuur).
Elke brug moet eenzelfde afstand overbruggen.
Leg uit waarom A en G altijd combineren met T, C en U
4.2 De genetische informatiestroom
We gaan hier niet het hele biologische proces behandelen, maar wel, in het kort, enkele scheikundige karakteristieken bij de vorming van de eiwitten in levende wezens.
Een samenvatting:
De informatiestroom gaat eerst van DNA naar RNA (transcriptie).
Het RNA, op zijn beurt, vertaalt haar codes voor het juiste eiwit (translatie)
DNA
(transcriptie)
RNA
(translatie / vertaling)
proteďnen
De basevolgorde van een gen (een deel van DNA of RNA) loopt parallel aan de aminozuurvolgorde van het product dat dat gen gaat maken (een polypeptide / eiwit).
De gencode ligt helemaal vast in de volgorde van de basen (A, C, T en U) en wel op zodanige manier dat elke drie opeenvolgende basen één aminozuur bepalen.
Dus (even vereenvoudigd): als een gen 600 basen heeft dan kan daaruit een eiwit van 200 aminozuren worden geproduceerd. Of eigenlijk moeten we hier niet het woord 'produceren' gebuiken, maar 'bepalen'.
Je zult begrijpen dat het van groot belang is om bij de juiste base te starten. Zo zijn er voor starten voor stoppen ook tripletcodes (codons).
Er zijn vier basen in het erfelijk materiaal, de genen, en daarmee kun je 43 verschillende aminozuren bepalen / definiëren. Maar je weet dat in levende wezens ongeveer 20 verschillende aminozuren worden gebruikt, geen 64.
Drie worden er gereserveerd voor het stoppen en starten van de productie, maar dan nog zijn er 61 over; nog steeds veel meer dan nodig.
In de praktijk kunnen meerder codes eenzelfde aminozuur bepalen en zulke codes (codons) worden ook wel synomieme codes genoemd.
Je weet dat het eiwitmolecuul (dus ook een enzym) gevormd wordt uit aminozuren in een polycondensatieproces.
De functie van proteďnen, in het bijzonder van enzymen, is buitengewoon belangrijk in het levende organisme, ook voor het metabolisme. Elk foutje in zo'n enzym kan tot gevolg hebben dat er stagnatie optreedt in het metabolisme. Dat kan ernstige (erfelijke) afwijkingen veroorzaken of zelfs het leven onmogelijk maken.
Dus is het controlemechaniek van het lichaam om toch vooral de juiste eiwitten / enzymen aan te maken zeer goed ontwikkeld. De informatie én de controle komt van het DNA, dat zich in de celkernen bevindt.
Dat DNA gaat niet zelf naar het plasma (de ribosomen) om die controle uit te voeren. Daarvoor stuurt hij kopiebrokken van zichzelf (RNA) (proces heet transcriptie) naar buiten, naar de ribosomen waar ook de aminozuren naar toe gesleept worden om daar in de juiste volgorde aan elkaar te worden gekoppeld.
Het volgende schema toont ongeveer zo'n gebeurtenis:
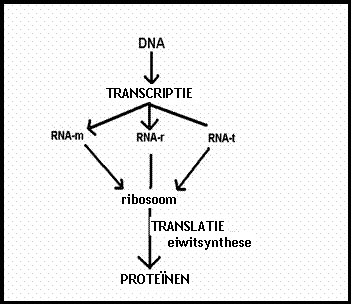
Bij de transcriptie worden delen van het DNA (genen) in de kern gekopieerd (met bijna dezelfde vier types nucleínezuren).
Bij de vertaling, de translatie, bij de ribosomen, worden de aminozuren (20 types in gebruik) aan elkaar gekoppeld tot eiwitten.
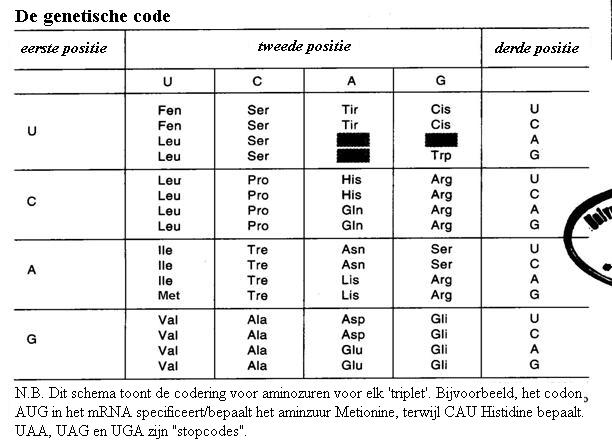
Opdracht 29
In de figuur van de dubbele helix van een DNA molecuul kunnen we in het onderste gedeelte een volgorde zien van de basen: - C - T - G - A - (van beneden naar boven).
T, na de transcriptie van DNA naar RNA, heeft die kleine wijziging ondergaan naar U, oftewel, dezelfde volgorde in RNA zal zijn: - C - U - G - A -
Welke aminozuren kunnen hier gevormd worden via dit stuk RNA? gebruik een gencode-tabel uit de biologie.
Kwantitatief gezien, dus in hoeveelheden, zijn er veel meer structuureiwitten in een levend wezen, dus eiwitten die de weefsels vormen: huid en haar, vlees, en dergelijke.
Echter, een ander type van eiwitten dat niet zozeer in grote hoeveelheden voorkomt, maar toch minstens zo belangrijk is, zo niet belangrijker, zijn de katalysatoren van de biochemische processen: de biokatalysatoren, de enzymen.
Een beetje geschiedenis
Louis Pasteur (1822 -1895) was een van de eerste wetenschappers die reacties onderzocht die gekatalyseerd worden door enzymen. Hij geloofde dat gist of levende bacteriën nodig waren voor zulke reacties, die hij "gisting" noemde. Bijvoorbeeld ook de omzetting van glucose in alcohol.
In 1897 maakte Eduard Büchner een filtraat zonder cellen dat afgezonderde enzymen bevatten. Daartoe behandelde hij gistcellen met fijn zand.
De enzymen die zich in dat filtraat bevonden waren prima in staat om glucose om te zetten in alcohol.
Zo bewees hij dat voor enzymactiviteit niet de aanwezigheid van levende cellen vereist is.
Büchner ontving hiervoor de Nobelprijs voor de scheikunde in 1907.
Twee algemene karakteristieken van enzymen:
- Het structuurdeel van enzymen is een eiwit. Opmerking hierbij: elke proteďnestructuur kan gedenatureerd worden, in het bijzonder onder invloed van pH en temperatuur.
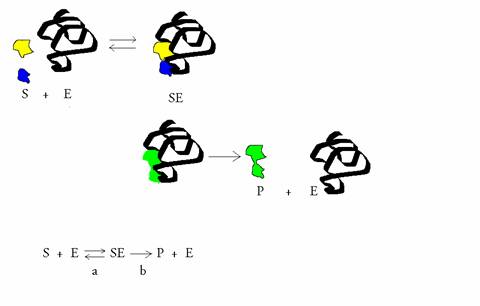
- Enzymen zijn (bio)katalysatoren en buitengewoon specifiek en efficiënt; het holo-enzym = apo-enzym + co-enzym/prostetische groep.
De werkzaamheid van biokatalysatoren hangt sterk af van de molecuulvorm, de ruimtelijke vorm, in het bijzonder het oppervlak van dat molecuul.
Bij voorbeeld: Stel je voor dat bepaalde moleculen van het substraat (de reagentia) willen reageren om een product te vormen. Dat zal alleen mogelijk zijn als die moleculen met elkaar in contact komen en dan nog zo dat ze op de juiste manier met elkaar in contact komen. (twee treinstellen koppelen ook alleen maar op één bepaalde manier aan elkaar).
Als die moleculen moeten wachten tot ze toevallig een keer botsen en dan ook nog op de juiste manier, dan kunnen ze lang wachten.
Maar dan komen de katalysatoren, de enzymen eraan om te helpen. Die halen moleculen naar zich toe, die passen in hun oppervlak en zo worden die moleculen precies op de goede manier naast elkaar gelegd om gemakkelijk en snel te reageren.
Als die moleculen eenmaal aan elkaar zitten, zal het enzym de boel weer loslaten en het nieuwe molecuul is gevormd.
Nogmaals: een substraat wil reageren. Het past precies in het oppervlak van het enzym, bij het actiecentrum. Eenmaal daar aangekomen kan het substraat prima reageren (met behulp van de plaatsen a, b en c).
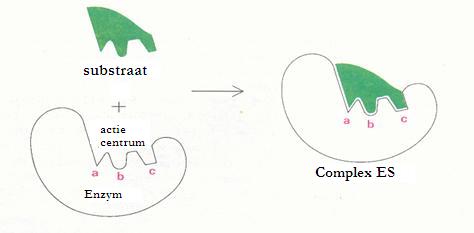
De reactievergelijking ziet er als volgt uit:
E + S ES E+P
S = substraat, P = product
Het functioneren van de enzymen heeft dus alles te maken met het oppervlak van haar moleculen.
Als iets of iemand dit oppervlak ook maar een beetje verandert, kan dat de werkzaamheid van het enzym lelijk verstoren.
Elk enzym heeft zijn temperatuur en pH waarbij de activiteit het grootst is: de optimale pH en temperatuur.
Opdracht 30
In het volgende schema zie je nóg een type van enzymkatalyse.
Bestudeer de afbeelding goed en verklaar hoe de specificiteit van dit enzym wordt gegarandeerd.
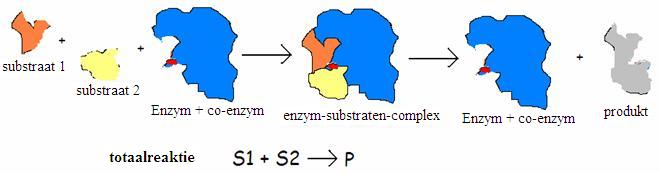
Opdracht 31
Kan de laatste reactie (S1+ S2 P) een condensatiereactie zijn?
Leg je antwoord uit.
Fouten in het DNA (mutaties) kunnen verwarring stichten in het (menselijk) lichaam. Misschien worden er dan foute enzymen aangemaakt, dus met een onjuiste ruimtelijke vorm.
Zulke foute enzymen maken de normale biochemische reactie in het metabolisme moelijk of zelfs onmogelijk. Zo iemand lijdt dan aan een ziekte.
Opdracht 32
Hemoglobine (Hb) mogen we beschouwen als een soort enzym met een activator. Welke activator heeft Hb?
Opdracht 33
Toon in een energiediagram wat "activeringsenergie" doet met een chemische reactie.
Antwoord 12-33
De twee substraten S1 en S2 verbinden zich heel specifiek aan het enzym. Andere moleculen passen gewoon niet (ruimtelijk gezien)
Regelmatig, maar niet in alle gevallen, bevindt zich in het actiecentrum van het enzym een co-enzym (zwak gebonden) of een prostetische groep (sterk gebonden).
Veranderingen van de vorm van het actiecentrum kan de enzymactiviteit sterk beďnvloeden, zelfs stil leggen.
Het centrum bevindt zich vaak in een soort spleet waarbinnen het vaak apolair is.
Coenzymen en prostetische groepen (  ) verbeteren de enzymactiviteit.
) verbeteren de enzymactiviteit.
Het complete enzym, inclusief de cofactoren en prostetische groepen, noemen we het holo-enzym,
Het enzym zonder cofactoren en prostetische groepen noemen we het apo-enzym.
Regelmatig is het co-enzym een vitamine, en hetzelfde co-enzym kan verschillende enzymen bedienen.
Sommige enzymen vereisen een anorganisch gedeelte, zoals een metaalionen (bijvoorbeeld, Ca2+, Mg2+ of Zn2+).
Deze anorganische component is een 'activator'. Funktioneel gezien is een activator gelijk aan een co-enzym, maar de anorganische componenten noemt met niet: co-enzymen.
Het enzym is - structureel gezien - een gewoon proteďne met:
- een primaire structuur, geproduceerd onder controle van DNA = de volgorde van de aminozuren
- een secundaire structuur, = de α-helix (schroef) van de primaire structuur
- een tertiaire structuur, = het resultaat als de α-helix zich opvouwt tot een driedimensionale vorm die de specificiteit van het enzym bepaalt evenals de manier waarop dit enzym geremd kan worden..
- een kwaternaire structuur (niet altijd aanwezig) is de samenvoeging van (vier) gelijke tertiaire structuren
verschillende soorten enzymen / nomenclatuur:
Meestal begint de naam van een enzym met het substraat + direct daarna de activiteit van dat enzym + uitgang -ase
| Oxireductases |
katalyseren redoxreacties (bijvoorbeeld:. glucose-oxidase) |
| Transferases |
transporteren functionele groepen van een gever naar een ontvanger. |
| hydrolases |
splitsen moleculen (voorbeeld: peptidases, proteases) |
| Liases |
onttrekken of voegen juist toe: bepaalde functionele groepen (voorbeeld. decarboxilase) |
| Isomerases |
veranderen isomeren (mutase) |
| Ligases |
verbinden verschillende moleculen met elkaar (piruvaatcarboxilase) |
Opdracht 34
Tot welk type behoren:
- Het enzym dat D-glucose kan veranderen in L-glucose?
- Het enzym dat fosfaat kan veranderen in fosfiet?
- Het enzym dat saccharose kan veranderen in glucose en fruktose?
Proteďnen, en zeker de enzymen, zijn zeer gevoelig voor het milieu waarin ze zich bevinden, met name w.b. de pH en de temperatuur. Die hebben grote invloed op de driedimensionale vorm.
Denaturatie bijvoorbeeld is desastreus: het enzym (een eiwit) verliest zijn tertiaire structuur en daarmee zijn ruimtelijke vorm. Dat enzym doet niets meer.
Dus wijk je af van de optimale pH en temperatuur, dan zal dat meteen de werkzaamheid van het enzym beďnvloeden.
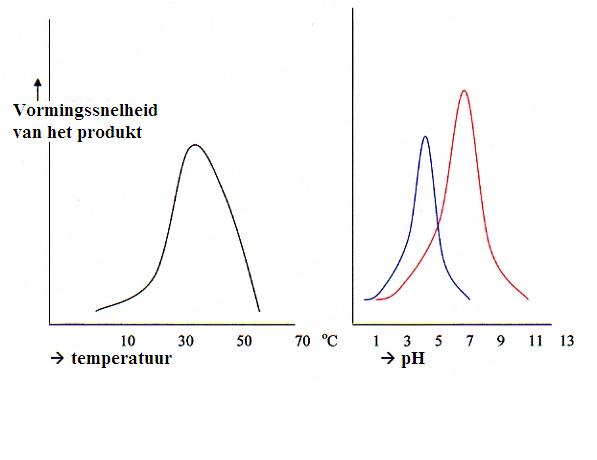
Een paar mogelijkheden om het milieu van het enzym te wijzigen:
- Verwarming geeft meer beweging van atomen, deeltjes in het algemeen en op bindingen. De toegenomen beweging kan vanderWaals bindingen en polaire aantrekkingen lelijk verstoren. Degelijke covalente bindingen gaan niet zo gemakkelijk stuk bij verwarming.
Als de denaturatie veroorzaakt werd door temperatuursverhoging is die verandering irreversibel (niet omkeerbaar). Zo'n enzym kun je weggooien. - Het toevoegen van zuur of alcohol kan de waterstofbruggen breken. Dan zal de tertiaire en misschien ook de secundaire structuur verloren gaan.
Je kunt zo de pH veranderen of op een andere manier het milieu anders maken door verdunnen of toevoegen van alcohol of zouten.
Het is mogelijk dat zulke denaturatie nog (gedeeltelijk) te herstellen is (reversibel).
In beide gevallen zal de tertiaire en misschien ook de secundaire structuur verloren gaan.
Opdracht 35
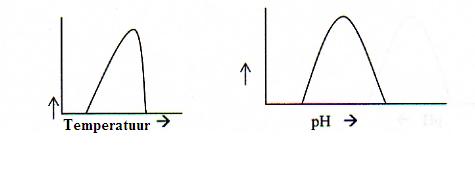
De figuren tonen de gevoeligheid van de enzymactiviteit voor temperatuur en pH. (de enzym-activiteit vind je op de y-as)
Maak in die figuur een mogelijke schaalverdeling van de x-as en de y-as (een schatting)
Opdracht 36
Leg uit wat er zal gebeuren bij het koken van een ei gedurende enkele minuten.
Hoge temperaturen zijn desastreus voor enzymen. Dit 'koken' is irreversibele denaturatie, onherstelbaar.
Voorzichtige en bescheiden temperatuursverlaging verlaagt alleen maar de activiteit, zonder dat er sprake is van denaturatie.
Ga je vanuit de verlaagde temperatuur terug naar de normale temperaturen van 30 - 40°C, dan komt de activiteit terug. (ideetje: voor je dood gaat je lichaam invriezen in de hoop dat het over honderd jaar weer tot leven kan worden gebracht)
Het merendeel van de enzymen functioneert het beste bij pH-waarden van ongeveer 7 (neutraal dus). Maar er zijn een paar uitzonderingen zoals pepsine, dat een optimale pH-waarde heeft bij 1 - 2.
Ook sterke pH schommelingen kunnen het enzym denatureren. Dat kan reversibel zijn, maar niet altijd.
Voorbeelden:
| Pepsine |
1,5 |
in de maag |
| Amilase |
6,6 |
in speeksel |
| Lipase |
8,0 resp. 7,0 |
in de pancreas resp. in de darmen |
| Saccharase |
7,0 |
in de maag |
Opdracht 37
Wat zal de pH in de mond zijn? Maak je antwoord aannemelijk.
Je weet dat de enzymactiviteit afhangt van het enzymoppervlak. Denaturatie houdt in dat de vorm, dus ook het oppervlak van het enzym verandert.
Opdracht 38
Tijdens de spijsvertering in het menselijk lichaam doen diverse enzymen mee, die elk zijn eigen pH-optimum heeft:
| waar: |
enzym |
pH-optimum |
| in het speeksel |
amilase e maltase |
6,6 |
| in de maag |
peptase, rennase |
1,5 - 4 |
| in de pancreas |
amilase, lipase, tryptase, polypeptidase |
6,6 - 9 |
| in de ingewanden |
maltase, saccharase, lactase, ereptase |
6,6 - 8,5 |
- Hoe zit dat met die pH-verschillen in deze vier plekken?
- Waarom verschijnt maltase pas helemaal aan het eind van het spijsverteringssysteem?
Antwoord 12-38
Niet gekatalyseerde reacties, die je bijvoorbeeld uren moet koken, zelfs al is er sterk zuur of base aanwezig, kunnen soms in een paar seconden bij fatsoenlijke temperatuur verlopen als het juiste enzym er bij zit, bij de optimale pH en temperatuur.
De enzymen treden op als katalysatoren door sterk de benodigde activeringsenergie te verlagen van deze specifieke biochemische reactie.
De verlaagde activeringsenergie staat toe dat zo'n proces dan verloopt op lichaamstemperatuur en met hoge snelheid
Er zijn diverse manieren om de enzymactiviteit te reguleren:
Verschillende enzymen staan niet meteen klaar om te functioneren, maar bevinden zich in een 'pre-enzym fase', of ook: de zymogene fase. Dit kan nodig zijn om het kwetsbare enzym te beschermen.
Het kan ook zijn dat het een enzym betreft dat pas nodig is in geval van ongelukken.
Zo hebben we in de maag pepsinogeen, dat het enzym pepsine kan vormen bij lage pH. Als het eenmaal is gevormd, kan het pepsine niet meer terug. Het pepsinogeen verliest een stuk keten van 44 aminozuren.
Het product zelf kan (als het hoge concentraties bereikt heeft) terugreageren en zo een enzym verbieden zijn activiteit voort te zetten.
Als het enzym bekend is kunnen specifieke stoffen worden toegevoegd die de enzymactiviteit beďnvloeden (remmers = moleculen met een structuur die lijkt op het normale substraat, maar toch niet helemaal gelijk is).
Zo maken die remmer het enzym (tijdelijk) inactief. Dus weer een vorm van regulering.
Eigenlijk hebben we het hier over drugs, geneesmiddelen die - bijvoorbeeld - gebruikt worden bij de chemotherapie door het metabolisme van tumoren te verbieden.
Sommige enzymen hebben naast een actiecentrum nóg een plek waar bepaalde groepen zich kunnen aanhechten: een zgn. allosterisch centrum.
Stoffen die in zo'n alosterisch centrum passen kunnen de driedimensionale vorm van het enzym wijzigen, inclusief het actiecentrum, en zo de specificiteit van dit enzym veranderen, en daarmee haar activiteit. Dat kan een positieve invloed zijn (actiever), maar ook een negatieve.
Opdracht 39
Zijn de volgende beweringen (a en b) waar of onwaar? Leg je antwoord uit.
- Een enzym is een (bio)katalysator die de reactiesnelheid beďnvloedt.
- Een katalysator versnelt (of vertraagt) de snelheid V van een chemische reactie: S P
- Geef een uitleg voor de letters S, P en V in je eigen woorden.
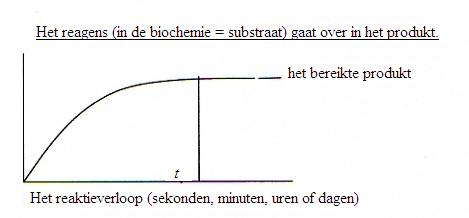
Meestal zijn biochemische reacties tevens evenwichtsreacties. Dat betekent dat de vorming van het product na enige tijd de maximum snelheid bereikt op het moment dat het evenwicht is bereikt.
Zonder katalysator (zie module 7) kan dat akelig lang duren: uren of zelfs dagen of helemaal nooit.
In aanwezigheid van een katalysator (een enzym) wordt dat evenwicht veel sneller bereikt. Dat wil dus niet zeggen dat je méér product krijgt, maar wel dat het sneller wordt gemaakt, mogelijk nu in een paar seconden.
Het enzym, de biokatalysator, versnelt meestal het chemisch proces met een factor 106 of meer.

Het substraat vormt een complex met een enzym. Dit complex wordt genoemd: de overgangstoestand. Het substraat verandert in product.
Vervolgens scheiden het product en het enzym zich van elkaar.
S + E ES P + E
a b
De vorming van het complex (a)is ook een evenwichtsreactie; de laatste reactie (b) is dat niet.
Het enzym versnelt het bereiken van het evenwicht, maar verandert het evenwicht zelf niet. Wel de voming van het product.
Elke reactie verloopt met zijn bepaalde reactiesnelheid V en met zijn eigen reactiesnelheidskonstante k
k1 k3
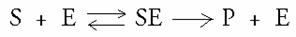
k2
Uitgaande van drie vooronderstellingen die een ideale / optimale situatie veronderstellen, mag je beweren dat de snelheid maximaal zal zijn,
Vmax

Die drie vooronderstellingen zijn:
- een stationaire toestand (steady state) waarin de concentratie van het intermediair ES niet verandert; de vorming van het ES complex vindt plaats met dezelfde snelheid als waarmee ES verdwijnt.
- Alle aanwezige enzym doet mee;
- Het systeem bevindt zich in een optimale omgeving (verzadigd met S, de beste pH, de beste temperatuur);
Op deze manier bereikt de snelheid van de reactie V haar maximale waarde Vmax
over de vormingssnelheid van het intermediair ES:

over de snelheid van het doorreageren / verdwijnen (in twee richtingen) van dat intermediair ES:


KM is de konstante van Michaelis
Opdracht 40
Leg uit hoe je aan die uitdrukking kan komen, uitgaande van de vooronderstelling 1
Antwoord 12-40
KM geeft de affiniteit van het enzym voor het substraat; een lage KM betekent veel ES, oftewel, veel substraat S,
en dat bekent: een lage KM wil zeggen: een grotere snelheid.
De verhouding tussen de reactiesnelheid en de KM kun je zien in de Michaelis Menten vergelijking:

De waarden van KM kunnen variëren van 0 tot duizenden.
Opdracht 41
Lees nauwkeurig de praktikumtaak en probeer deze beschrijving te begrijpen en van je commentaar te voorzien.:
“Je verdeelt een neutraal speekselextract (pH=7) in twee gelijke delen (A en B).
Deel A wordt aangezuurd met HCl totdat de pH=2. Dit mengsel toont geen activiteit meer.
Deel B wordt vermengd met een gelijk volume water. Dit mengsel vertoont geen activiteit meer.
Daarna behandelen we de beide oplossingen, A en B met base totdat de pH=8,5. Er is geen enzymactiviteit meer.
Tenslotte worden de twee delen met pH 8,5 aangezuurd tot pH=2. Alleen deel B toont weer activiteit.”
Opdracht 42
Probeer de volgende proefbeschrijving te begrijpen, mede gebruik makende van grafieken:
"Bij vergelijking van de enzymen bepalen we de parameters Vmax en Km en meten we de reactiesnelheid (V) als functie van de substraatconcentratie [S]."
Voor een bepaalde standaardhoeveelheid hexokinase of glycokinase meten we de vormingssnelheid (V) van glucosefosfaat (het product G6P) als functie van de glucoseconcentratie [S].
De resultaten tonen aan dat Vmax van de twee enzymen aan elkaar gelijk zijn (er vormen zich ±100 nmol G6P per minuut), maar dat de Km-waarden flink verschillen:
Km van hexokinase = 0,1 mM, Km da glucokinase = 10 nM (factor 100)."
Kijk nog eens goed naar het volgende evenwicht met de tussenstof SE:
k1
k3
k2
De Michaelisconstante
 geeft de affiniteit van het enzym voor het substraat.
geeft de affiniteit van het enzym voor het substraat.
De vergelijking van Michaelis-Menten
geeft de relatie tussen KM en de reactiesnelheid.
We kennen diverse types van enzymremmers:
- productremmers
- drugsremmers
- allosterische remmers
http://www.le.ac.uk/by/teach/biochemweb/tutorials/michment2print.html
Enzyme kinetics and the Michaelis-Menten equation
Introduction
Study of the impact made on the rate of an enzyme-catalysed reaction by changes in experimental conditions is known as enzyme kinetics.
Knowledge of kinetics can be a very useful tool in understanding the mechanism by which an enzyme carries out its catalytic activity.
The effect of substrate concentration on the initial rate of an enzyme-catalysed reaction is a central concept in enzyme kinetics. When data are generated from experiments of this type and the results plotted as a graph of initial rate (v, y-axis) against substrate concentration ([S] *, x-axis), many enzymes exhibit a rectangular hyperbolic curve like the one shown in the diagram below.
*Note: the use of square brackets, as for [S] above is short-hand notation for "concentration of S", a convention that will be used extensively in the derivation below.

Observations of this type set Leonor Michaelis and Maud Menten thinking about the underlying reasons why a curve should follow this shape and led them to derive an algebraic equation that now bears their names. There are several modern ways to explain the way in which the Michaelis-Menten equation is derived, and one is spelt out below.
Deriving the Michaelis-Menten Equation
Start with the generalised scheme for enzyme-catalysed production of a product (P) from substrate (S). The enzyme (E) does not magically convert S into P, it must first come into physical contact with it, i.e. E binds S to form an enzyme-substrate complex (ES).
Michaelis and Menten therefore set out the following scheme:
The terms k1, k-1 and k2 are rate constants for, respectively, the association of substrate and enzyme, the dissociation of unaltered substrate from the enzyme and the dissociation of product (= altered substrate) from the enzyme. Note that there is the theoretical possibility of a reverse reaction, with ES complex forming from E and P, but this can be ignored because we are considering initial rates of reaction, i.e. when the enzyme is first provided with substrate, so there should not be any product available to combine with enzyme.
The overall rate of the reaction (v) is limited by the step ES to E + P, and this will depend on two factors - the rate of that step (i.e. k2) and the concentration of enzyme that has substrate bound, i.e. [ES]. This can be written as:
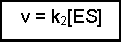 (Equation 1)
(Equation 1)
At this point it is important to draw your attention to two assumptions that are made in this scheme. The first is the availability of a vast excess of substrate, so that [S]>>[E]. Secondly, it is assumed that the system is in steady-state, i.e. that the ES complex is being formed and broken down at the same rate, so that overall [ES] is constant. The formation of ES will depend on the rate constant K1 and the availability of enzyme and substrate, i.e. [E] and [S]. The breakdown of [ES] can occur in two ways, either the conversion of substrate to product or the non-reactive dissociation of substrate from the complex. In both instances the [ES] will be significant. Thus, at steady state we can write:
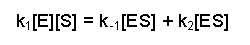
The next couple of steps are rearrangements of this equation. First of all we can collect together the rate constants on the right-hand side because they are both multiplied by [ES], this gives us:
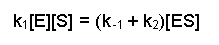
Then dividing both sides by (k-1 + k2), this becomes:
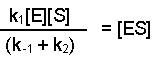
Notice that the three rate constants are now on the same side of the equation. As the name implies, these terms are constants, so we can actually combine them into one term. This new constant is termed the Michaelis constant and is written KM.
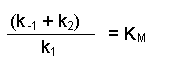
Notice that the three rate constants in the definition of KM are actually inverted (the other way up) compared with our previous equation. This is a 'trick' that makes for easier calculation at a later stage. Substituting this definition of KM into our previous equation now gives us:
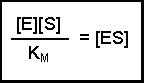 (Equation 2)
(Equation 2)
The total amount of enzyme in the system must be the same throughout the experiment, but it can either be free (unbound) E or in complex with substrate, ES. If we term the total enzyme E0, this relationship can be written out:
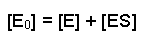
This can be rearranged (by subtracting [ES] from each side) to give:
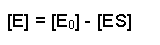
So, the [E] free in solution is equal to the total amount of enzyme minus the amount that has substrate bound. Substituting this definition of [E] back into equation 2 gives us:
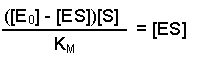
This can now be rearranged in several steps. First of all, open the bracket so that the terms [E0] and [ES] are separately multiplied by [S]
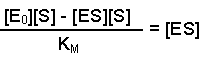
Next, multiply each side by KM, this gives us:

Then collect the two [ES] terms together on the same side (you can either think of this as adding [ES][S] to both sides or as 'carry over and change the sign' - your preference will probably be an indication of how long ago you went to school). This gives:
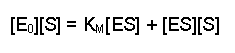
Then because both terms on the right-hand side are multiplied by [ES] we can collect them together into a bracket:
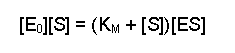
Dividing both sides by (KM + [S]) now gives us:
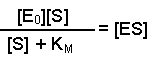
Substituting this left-hand side into Equation 1 in place of [ES] results in:
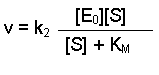
The maximum rate, which we can call Vmax, would be achieved when all of the enzyme molecules have substrate bound. Under conditions when [S] is much greater than [E], it is fair to assume that all E will be in the form ES. Therefore [E0] = [ES]. Thinking again about Equation 1, we could substitute the term Vmax for v and [E0] for [ES]. This would give us:
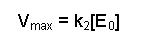
Notice that k2[E0] was present in our previous equation, so we can replace this with Vmax, giving a final equation:
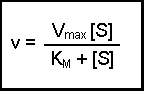
This final equation is actually called the Michaelis-Menten equation.
So what?
Perhaps this derivation still leaves you puzzled about the importance of the Michaelis-Menten equation. The significance becomes clearer when you consider the case when the rate of reaction (v) is exactly half of the maximal reaction rate (Vmax). Under those circumstances, the Michaelis-Menten equation could be written:
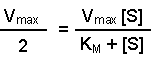
On dividing both sides by Vmax this becomes:
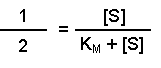
Multiplying both sides by (KM + [S]) gives:
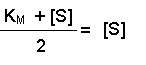
And then multiplying both sides by 2 further resolves the equation to:
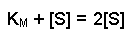
2[S] on the right-hand side is the same as [S] + [S], so we can take away one [S] from each side. Thus when the rate of the reaction is half of the maximum rate:
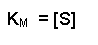
The KM of an enzyme is therefore the substrate concentration at which the reaction occurs at half of the maximum rate. If we now reconsider the graph that came at the start of this tutorial it could be written:
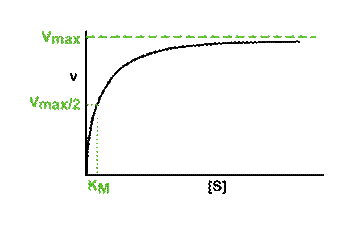
What does this all mean in physical terms? KM is an indicator of the affinity that an enzyme has for a given substrate, and hence the stability of the enzyme-substrate complex.
Look at the shape of the graph. At low [S], it is the availability of substrate that is the limiting factor. Therefore as more substrate is added there is a rapid increase in the initial rate of the reaction - any substrate is rapidly mopped up and converted to product. At the KM, 50% of active sites have substrate bound. At higher [S] a point is reached (at least theoretically) where all of the enzyme has substrate bound and is working flat out. Adding more substrate will not increase the rate of the reaction, hence the levelling out observed in the graph.
There are limitations in the quantitative (i.e. numerical) interpretation of this type of graph, known as a Michaelis plot. The Vmax is never really reached and therefore Vmax and hence KM values calculated from this graph are somewhat approximate. A more accurate way to determine Vmax and KM (though still not perfect) is to convert the data into a linear Lineweaver-Burk plot. See separate page:
"Experimental calculation of Vmax and KM "
If, incidentally, we consider that the rate constant k2 for the conversion of ES to E + P in the initial scheme is the step determining the overall rate of production of P (as we have in deriving the Michaelis-Menten equation) then k2 is actually the same term as kcat, the turnover number.